|
|||||||
| Δίσκοι Κατά πολλούς οι βασιλιάδες του ενυδρείου. |
 |
|
|
Εργαλεία Θεμάτων | Τρόποι εμφάνισης |
|
#1
 10-03-14, 22:52
10-03-14, 22:52
|
||||
|
||||
|
Το περιβάλλον που ζουν οι δίσκοι στη φύση 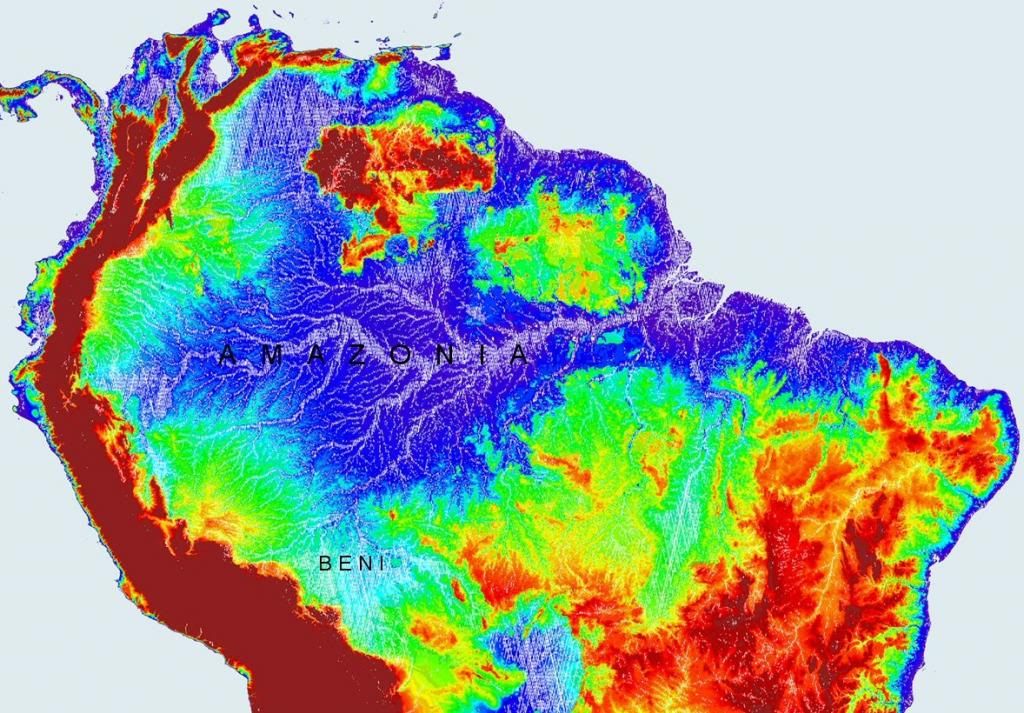 Με το μπλε χρώμα φαίνεται η έκταση των πλημμυρών κατά τη διάρκεια της εποχής των βροχοπτώσεων.
Οι δίσκοι είναι επίσης μοναδικοί ανάμεσα στις κιχλίδες που σχηματίζουν μεγάλα στατικά κοπάδια που αποτελούνται από εκατοντάδες ψάρια κατά τη διάρκεια της ξηρής περιόδου. Οι τοπικοί ψαράδες τα ονομάζουν αποικίες. Αυτές οι αποικίες βρίσκουν καταφύγιο σε χοντρές ρίζες και κλαδιά που έχουν πέσει στη λίμνη που οι ντόπιοι τα ονομάζουν galhadas. Στη περίοδο πλημμυρών διασκορπίζονται στα πλημμυρισμένα δάση. Η μελέτη πραγματοποιήθηκε σε δύο λίμνες με blackwater στο Uxi Bay κοντά στο lago Amana και σε μια κοντινή περιοχή στο lago Urini. Επελέγησαν οι περιοχές αυτές γιατί δεν υπήρχε επαγγελματικό ψάρεμα δίσκων στο παρελθόν και επίσης γιατί εκεί οι πληθυσμοί των πιράνχας (Pygocentrus nattereri - τα γνωστά σε μας red belly και Serrasalmus spp) σε σχέση με τους whitewater ποταμούς ήταν μικροί. Παράλληλες μελέτες σε άλλες περιοχές του Αμαζονίου και στον ποταμό Japura εγκαταλείφθηκαν καθώς οι ζημιές στα δίχτυα από τα πιράνχας ήταν πολύ σοβαρές. Το Uxi Bay επιλέχθηκε γιατί είχε μικρό πληθυσμό δίσκων που επέτρεψε να προλάβουν να επεξεργαστούν όλο το πληθυσμό των δίσκων πριν αλλάξει η στάθμη του νερού. Είναι επίσης απομονωμένοι από άλλους πληθυσμούς δίσκων εξαιτίας μιας τουλάχιστον 6 χιλιομέτρων, ρηχής και με κύματα ακτογραμμής (βιότοπος που αποφεύγουν οι δίσκοι). Εκεί υπήρχε μεγαλύτερος αριθμός δίσκων αλλά δεν υπήρχε χώρος να τους κρατήσουν όλους κάνοντας αδύνατη την μέτρηση όλων των ψαριών. Ο χρωματικός φαινότυπος των δίσκων στις παραπάνω περιοχές ήταν ο μπλε/καφέ - Symphysodon haraldi (κατά Bleher).  Στην φωτογραφία από δορυφόρο διακρίνουμε την ίδια περιοχή σε περίοδο βροχοπτώσεων (πάνω) και σε περίοδο "ξηρασίας" (κάτω) Η ζωή όλων των οργανισμών που ζουν στο μεγαλύτερο μέρος του Αμαζονίου επηρεάζεται από τον ατέρμονο κύκλο της περιόδου των βροχοπτώσεων και της εναλλαγής της με μιας πιο ξηρής περιόδου. Οι συνεχιζόμενες βροχοπτώσεις προκαλούν εκτεταμένες πλημμύρες ενώ αντίθετα η έλλειψη βροχοπτώσεων προκαλεί σημαντική μείωση της στάθμης του ποταμού. Σε όλο το μήκος του ποταμού αυτές οι περίοδοι ξεκινούν και σταματούν σε διαφορετικούς μήνες. Πολλά παρακλάδια του Αμαζονίου πλημμυρίζουν τον Νοέμβρη και συνεχίζει να ανεβαίνει η στάθμη μέχρι τον Ιούνιο. Ο Rio Negro ξεκινάει να πλημμυρίζει Φεβρουάριο με Μάρτιο ενώ ο Madeira 2 μήνες πιο πριν! Η εν λόγω μελέτη έγινε στο Tefe της Βραζιλίας. Η περίοδος βροχοπτώσεων και εκεί ξεκινάει από τον Φεβρουάριο (300-450 mm/μήνα) και από τον Αύγουστο πιο "ξηρής" περιόδου με 50-150 mm/μήνα. Η άνοδος της στάθμης φτάνει από τα 8 έως τα 16 μέτρα με αποκορύφωμα κάθε χρόνο τον Απρίλιο-Ιούνιο και με τις χαμηλότερες στάθμες τον Οκτώβριο. Παράμετροι νερού   Φωτογραφία 2: Από δορυφόρο διακρίνουμε τον blackwater και τον whitewater ποταμό Φωτογραφία 3: Η ίδια περιοχή πλημμυρισμένη 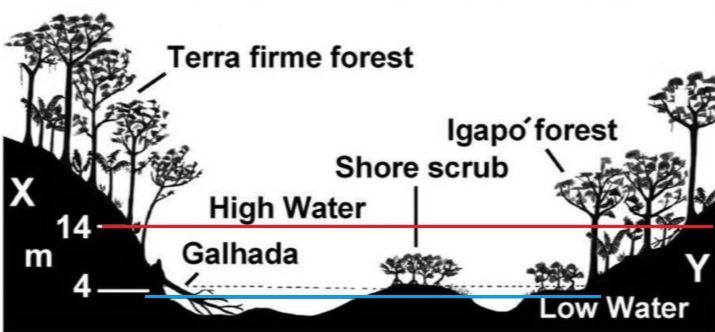 Διακρίνουμε τη στάθμη του νερού κατά τη διάρκεια της ξηρής περιόδου (μπλε γραμμή) και πόσο ανεβαίνει τη περίοδο των βροχοπτώσεων (κόκκινη γραμμή). Επίσης διακρίνουμε και τα ονομαζόμενα galhadas - στην ουσία είναι δέντρα που έχουν πέσει στο νερό. Δειγματοληψία Για να μεγιστοποιηθεί το ποσοστό των δίσκων που βρίσκονται σε κατάσταση αναπαραγωγής, η δειγματοληψία έγινε κατά την έναρξη της σεζόν της αύξησης της στάθμης του νερού(Δεκέμβριος 1997 και Νοέμβριος του 1998 για το Uxi Bay και Δεκέμβριος 1999 στην λίμνη Urini) πριν πλημμυρίσουν τα galhadas και δεν είναι αποτελεσματική η δειγματοληψία λόγω του μεγάλου βάθους. Επιπλέον δείγματα ελήφθησαν στη χαμηλότερη άμπωτη τον Οκτώβριο του 1997 στο Lago Urini. Με την βοήθεια μιας ομάδας από 8-10 ψαράδες κάθε galhada περικυκλώθηκε με δίχτυ 50 x 8 m (με πλέγμα 5 mm), αφαιρέθηκαν τα κλαδιά με τσεκούρια και πριόνια. Ο αριθμός των ψαριών ανά galhada είναι μια σχετική εκτίμηση επειδή μερικά ψάρια διέφυγαν γύρω ή πάνω από το δίχτυ. Μετά την καταμέτρηση του πληθυσμού κάθε galhada ανακατασκευάστηκε και προστέθηκαν φρεσκοκομμένα κλαδιά μήκους 2-4 m. Μία εβδομάδα πριν την δειγματοληψία στο Uxi Bay έφτιαξαν 25 "τεχνητά" galhadas από τους σωρούς κλαδιών σε σημεία μεταξύ των υφιστάμενων φυσικών για να εξετάσουν αν θα επιλεγούν από δίσκους και άλλες κιχλίδες. Έγινε δειγματοληψία από παραλίες, από περιοχές που φύονταν ομάδες από Cyperus. Επίσης σε πλημμυρισμένες ακτές και στα "ανοιχτά". Για να πιάσουν και παρατηρήσουν τους δίσκους την νύχτα χρησιμοποιήθηκαν φακοί και δίχτυα. Κατά τη διάρκεια της εποχής των πλημμυρών περίπου 100 ώρες παρατήρησης έγιναν από το κανό σε πλημμυρισμένα δάση της λίμνης Amana.  Φωτογραφία από την σύλληψη δίσκων Οι δίσκοι κρατήθηκαν σε δεξαμενές με αεραντλίες σε μια βάρκα. Μετρήθηκε το μήκος (standard length - SL) σε mm και το βάρος σε gr. Επίσης μετρήθηκαν τα παράσιτα στα βράγχια που είχε το κάθε ψάρι (πρόκειται για τα Cymothoidae). Μερικά ψάρια από τα 218 ψάρια από το Uxi Bay θυσιάστηκαν χωρίς να υποφέρουν και εξετάστηκε η ανατομία τους ώστε να καθοριστεί το φύλο τους και η ωριμότητα τους.    Symphysodon haraldi (κλικ για μεγέθυνση) Τον Δεκέμβριο του 1997 σε 91 δίσκους τοποθετήθηκαν ταμπελάκια (tags) και στη συνέχεια απελευθερώθηκαν στο ίδιο μέρος που πιάστηκαν (στο Uxi Bay) με τη προσδοκία ότι θα συλληφθούν ξανά το επόμενο έτος και θα διαπιστωθεί η ανάπτυξη τους αλλά και αν βρεθούν στα ίδια μέρη. Επίσης στις 16-18 Νοεμβρίου 1998 τοποθετήθηκαν ταμπελάκια (tags) σε 219 δίσκους και απελευθερώθηκαν εντός 12 ωρών στο Uxi Bay ., 1998. Μετά από 7-9 ημέρες αργότερα σε όλα τα galhadas του Uxi Bay έγινε ξανά δειγματοληψία ώστε να τεκμηριωθούν οι βραχυπρόθεσμες διασπορές του πληθυσμού. Οι ετικέτες ήταν αριθμημένες με διαστάσεις 13 χ 2 mm και τοποθετήθηκαν στη βάση του ραχιαίου πτερυγίου κάθε δίσκου. Δοκιμάστηκαν πρώτα σε 15 δίσκους 100-150 χιλιοστά SL σε ένα ενυδρείο για τρεις εβδομάδες. Όλα έμειναν στη θέση τους και κανένα από τα ψάρια δε μολύνθηκε. Ανάλυση περιεχομένου στομαχιών των δίσκων Για να δοθεί έμφαση στην επίδραση της κάθε εποχής (ξηρασίας και πλημμύρας) στις διατροφικές τους συνήθειες οι δίσκοι συλλέχθηκαν κατά το τέλος της χαμηλής περιόδου (ξηρασίας) και κατά την αποκορύφωση της υψηλής σεζόν (πλημμύρας). Τα στομάχια τους διατηρήθηκαν σε διάλυμα 10% φορμαλδεΰδης πριν την εργαστηριακή ανάλυση. Αντίκτυπος του συνεχόμενου μεταβαλλόμενου περιβάλλοντος στη ζωή των δίσκων Είναι προφανές ότι η σύσταση του νερού ακόμα και το περιβάλλον που ζουν οι δίσκοι αλλάζουν συνεχώς. Πρόκειται για το μεγαλύτερο και παραγωγικό οικοσύστημα σε ολόκληρη τη γη και εξαιρετικά πλούσιο σε είδη ζωντανών οργανισμών. Η ετήσια αυξομείωση της στάθμης εκθέτει όλους τους οργανισμούς που ζουν στον Αμαζόνιο σε μεγάλες διακυμάνσεις από την διαθεσιμότητα τροφής και προστασίας από θηρευτές, την πυκνότητα των παρασίτων και των θηρευτών έως τις φυσικοχημικές ιδιότητες του νερού όπως πχ το διαθέσιμο οξυγόνο. Οι δίσκοι όπως και όλα τα ψάρια στον Αμαζόνιο το καθένα με τον τρόπο του έπρεπε να προσαρμοστούν για να επιβιώσουν. Θα είναι πολύ χρήσιμο να τα εξετάσουμε τι λύσεις "βρήκαν" οι δίσκοι σε όλα αυτά. Ανάμεσα σε αυτές είναι η καλύτερη κατανόηση της προσαρμογής στην υποξία που υπόκεινται (Val & Almeida- Val, 1995). Επίσης η δίαιτα και οι πηγές ενέργειας κατά τον κύκλο της ξηρασίας-πλημμύρας σχετικά είναι γνωστή (Araujo- Lima et al., 1986; Goulding et al., 1988; Benedito-Cecilio et al., 2000). Ομοίως η κατανομή και οι κινήσεις των ψαριών ανάλογα με τις μεταβολές στο οξυγόνο και τη διαθεσιμότητα της τροφής είναι επαρκώς τεκμηριωμένες από προηγούμενες μελέτες (Goulding, 1980, Cox-Fernandes, 1997, Crampton, 1998, Henderson et al., 1998, Petry et al., 2003, Granado-Lorencio et al., 2005, Correa et al. 2008 ) 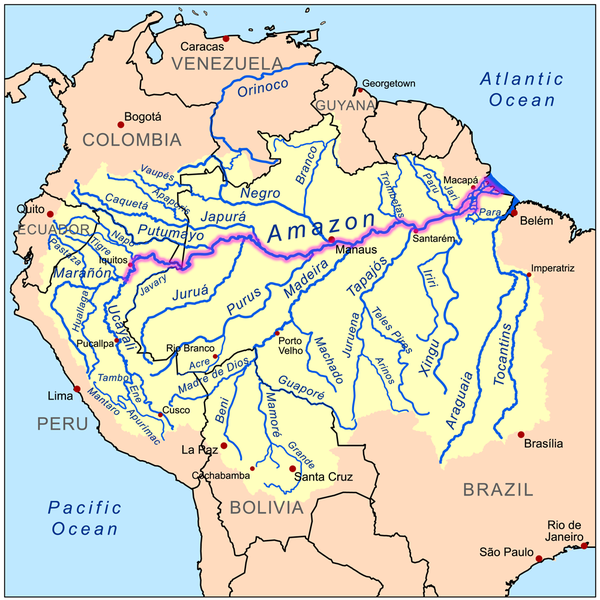 Χάρτης που δείχνει το τεράστιο μέγεθος του Αμαζονίου. (κλικ για μεγέθυνση) Αποτελέσματα: Επίπεδο πλημμύρας και ηλεκτρική αγωγιμότητα. Η ηλεκτρική αγωγιμότητα του νερού στο Uxi bay παρουσίασε μια αντίστροφη σχέση με το επίπεδο του νερού, κυμαινόμενη από 8 mScm-1 σε περίοδο πλημμύρας σε 29 mScm-1 σε περίοδο ξηρασίας. Κατά τη διάρκεια των δύο ετών της μελέτης καθώς επίσης και κατά τα τρία προηγούμενα έτη κατά τη διάρκεια των οποίων η στάθμη του νερού καταγράφονταν, γεγονότα που είναι γνωστά στους ντόπιους ως «Repiquetes» κατά τη διάρκεια κάθε πλημμύρας. Στην ουσία πρόκειται για περιόδους που η στάθμη του νερού πέφτει η μένει σταθερή με συνέπεια να διακόπτεται η ανοδική τάση των νερών της πλημμύρας. Τα εν λόγω Repiquetes συνδέονται με τις διακυμάνσεις στην ηλεκτρική αγωγιμότητα. 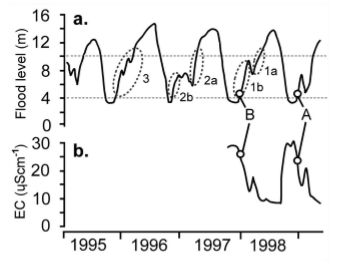 (a) το επίπεδο των πλημμυρών και (b) ηλεκτρική αγωγιμότητα (EC) που παρατηρήθηκαν στον Lago Amana κατά τη διάρκεια της περιόδου 1995-1998. Οι διακεκομμένοι κύκλοι αντιπροσωπεύουν τις περιόδους που γεννούν οι δίσκοι Διατροφή των δίσκων. Η μέση πληρότητα του στομάχου, κατά την ξηρασία ήταν 28,4% με το 33% των δίσκων να έχουν εντελώς άδειο στομάχι και το 68% να έχει πληρότητα 30% ή λιγότερο. Σε περιόδους πλημμύρας η μέση πληρότητα του στομάχου ήταν 89,2% με κανένα ψάρι να εμφανίζει πληρότητα του στομάχου μικρότερη από 50%. Η αναλογική σύνθεση των ειδών διατροφής που βρέθηκαν στα στομάχια των δίσκων παρουσιάζονται στο διάγραμμα που ακολουθεί: 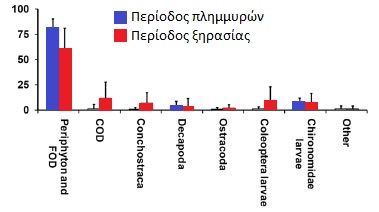 Οι αλλαγές του τύπου της διατροφής των δίσκων σε σχέση με την περίοδο που βρέθηκαν σε στομάχια δίσκων από blackwater βιότοπο στην περιοχή Tefé. Periphyton = Άλγη FOD = Fine organic detritus (οργανική ύλη) GPM = green plant (macrophytes) matter – ως επί το πλείστον μικρά κονιοποιημένα κομμάτια φύλλων. COD = Coarse organic detritus – κυρίως κομμάτια ξύλου και φλοιού. Conchostraca Decapoda Ostracoda Coleoptera larvae Chironomidae larvae Τα Periphyton, FOD and GPM ήταν δύσκολο να υπολογισθούν ποσοτικά και μπήκαν στην ίδια κατηγορία. GPM ήταν μόνο περίπου το 15% αυτής της κατηγορίας. Προτίμηση μικροενδιαιτήματων και κατανομή των δίσκων μεταξύ των galhadas. Το1997 το 97,8% των δίσκων που πιάστηκαν και το 1998 αντίστοιχα το 86,8% βρίσκονταν σε ένα μεγάλο galhada στην πιο βαθιά περιοχή του κόλπου (2-3 μ.), είχε σχεδόν καθόλου ροή (<0,01 ms-1) και καλά προστατευμένη από τα κύματα. Έξω από το μεγάλο αυτό galhada οι υπόλοιποι δίσκοι βρέθηκαν μόνο σε galhadas με διάμετρο μεγαλύτερη από 4 m που βρίσκονται σε σχεδόν στάσιμο νερό σε βάθος 1,5 έως 3,0 μέτρα και προστατευμένα από τα κύματα. Οι ερευνητές κατέγραψαν επίσης συναθροίσεις δίσκων που αριθμούν σε αρκετές δεκάδες ψάρια σε βαθιές (1-2,5 m) λίμνες του που δημιουργούνται από ένα παραπόταμο που ρέει μέσα στο τροπικό δάσος Uxi Bay (J. de Oliveira Alves, άτομα. comm.). Στο lago Urini ερευνήθηκαν συνολικά 70 galhadas . Όλα ήταν μεγαλύτερα από 4 m σε διάμετρο που βρίσκονται σε περιοχές προστατευμένες από την ισχυρή δράση των κυμάτων και βρίσκονταν σε σημεία με αμελητέα ροή του νερού (<0,01 ms-1). Δεν υπήρχαν δίσκοι σε 48 από αυτά τα galhadas ενώ ήταν παρών σε 22. Χωρίσαμε αυτά τα 70 galhadas σε τρεις αυθαίρετες κατηγορίες. Η πρώτη περιελάμβανε εκείνες σε νερό με μικρότερο από 1 m βάθος (στο χαμηλότερο επίπεδο του νερού), η δεύτερη σε νερό με 1-2 m βάθος και η τρίτη σε νερό 2-4 m βάθος. Οι δίσκοι απουσίαζαν από την πρώτη κατηγορία. Η δεύτερη κατηγορία βάθος περιείχε 24 δίσκους (1,9% των συνολικών ψαριών που βρέθηκαν) ενώ στη τρίτη κατηγορία βρέθηκαν 1.246 δίσκοι (98,1%). Οι ντόπιοι ανέφεραν ότι σε galhadas που βρίσκονται σε παρόμοια βάθη σε άλλα τμήματα της λίμνης ότι φιλοξενούν περισσότερους από χίλιους δίσκους. Μόνο 16 από τα 41 galhadas στη κατηγορία βάθους 2-4 m ( 39 % ) είχαν καταληφθεί από δίσκους. Τα υπόλοιπα «κενά» από δίσκους galhadas δεν διαφέρουν από αυτά που φιλοξενούσαν δίσκους όσον αφορά τη δομή τους (διακλάδωση), τα ιζήματα , τη σκίαση ή από τον βαθμό στον οποίο προστατεύονται από τον άνεμο και τη δράση των κυμάτων. Σε περιόδους πλημμύρας οι αποικίες των δίσκων διασκορπίζονται μέσα σε πλημμυρισμένα δάση και πυκνές θαμνώδες ακτές. Τα galhadas που φιλοξενούσαν τους περισσότερους δίσκους στο Uxi Bay ερευνήθηκαν την 1η Δεκεμβρίου 1998 μετά από άνοδο της στάθμης των υδάτων (2,5 m). Δεν βρέθηκε κανένας δίσκος. Κατά τη διάρκεια νυχτερινής παρατήρησης σε πλημμυρισμένα δάση κατά τη διάρκεια των πλημμυρών οι δίσκοι ήταν συχνά μόνοι τους ή σε ομάδες έως έξι ψαριών αλλά ποτέ σε μεγαλύτερες ομάδες (των 6). Κατά τη διάρκεια της αποκορύφωσης της πλημμύρας το 82 % (237 από τους 289) των δίσκων παρατηρήθηκαν σε πυκνές θαμνώδες ακτή ή σε δάση με δέντρα μέτριου ύψους ( < 15 m) . Το υπόλοιπο 18 % βρέθηκε σε περισσότερα σκιασμένα δάση του Igapó που έχει πιο ψηλά δέντρα (15 - 25 m).  Εκεί που ο whitewater συναντά τον blackwater Περιοχή και μετακίνηση δίσκων. Από τους 91 δίσκους που είχαν τοποθετήσει ετικέτες (tags) στο Uxi Bay το Δεκέμβριο του 1997, μόνο ένας αρσενικός δίσκος βρέθηκε το Νοέμβριο του 1998. Πιάστηκε ξανά στην ίδια ακριβώς θέση 11 μήνες αργότερα. Κατά τη διάρκεια αυτής της περιόδου ο δίσκος μεγάλωσε από 108 σε137 mm SL και από 71 σε 151 gr σε βάρος. Αυτό μας δίνει πολλές πληροφορίες για την ετήσια ανάπτυξη των δίσκων. 7-9 ημέρες αργότερα βρέθηκαν 104 από τους 219 δίσκους που είχαν ετικέτα ενώ ερευνήθηκαν όλα τα 58 φυσικά και τεχνητά galhadas στον κόλπο. Οι αποστάσεις της ακτογραμμής που διανύονται από τα ψάρια κατά τη διάρκεια αυτής της περιόδου δείχνουν μικρή βραχυπρόθεσμη διασπορά, παρά το γεγονός ότι το άγχος της σύλληψης και τοποθέτησης ετικετών συχνά προκαλεί την μετακίνηση των ζώων (Southwood & Henderson, 2000). Τα 25 τεχνητά galhadas που φτιάχτηκαν μία εβδομάδα πριν τη δειγματοληψία του 1998 αποικίστηκαν από 111 κιχλίδες αλλά μόνο επτά (6,3%) από αυτές ήταν δίσκοι. 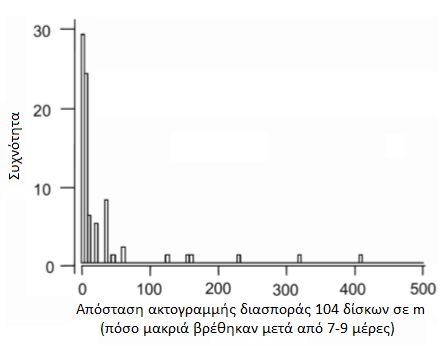 Κατά τη διάρκεια αυτής της μελέτης ένας μικρός δίσκος (75 χιλιοστά SL) βρέθηκε στο στομάχι ενός Serrasalmus rhombeus 1,1 κιλών αλλά σε κανένα από τα ακόλουθa είδη που αναφέρουν οι ντόπιοι ότι τρέφονται με δίσκους: Serrasalmus spp, Pygocentrus nattereri, Hoplias malabaricus, Hydrolycus spp, Electrophorus electricus , Pseudoplatystoma fasciatum, Ν. tigrinum (Valenciennes), Plagioscion squamosissimus (Heckel) και Cichla monoculus.  Παρασιτισμός. 69 από τους 492 δίσκους (14%) είχαν Β. cichlae. Εφτά ψάρια είχαν δύο παράσιτα (B. cichlae) (ένα σε κάθε βραγχιακό θάλαμο) και τα υπόλοιπα 62 είχαν μόνο ένα στον βραγχιακό θάλαμο. Κανένας από τα δίσκους που είχε προσβληθεί δεν είχε προχωρήσει σε σεξουαλική ωριμότητα. Επίσης οι δίσκοι αυτοί είχαν σημαντικά λιγότερο βάρος.  Παράσιτα δίσκων στη φύση (κλικ για μεγέθυνση) Φύλο και αναπαραγωγική ετοιμότητα. 218 δίσκοι από το Uxi Bay υποβλήθηκαν σε ανατομή και καθορίστηκε το φύλο τους. 71 ήταν αρσενικά και 68 θηλυκά . Οι υπόλοιποι 79 ήταν ανήλικοι απροσδιόριστου φύλου. Η σχέση μεταξύ του SL (Standard length) και του βάρος για τoυς αρσενικούς και θηλυκούς δίσκους δεν ήταν σημαντικά διαφορετική μεταξύ των δύο φύλων. Παρόλα αυτά τα αρσενικά είχαν μεγαλύτερο μέγεθος από ότι τα θηλυκά (152 χιλιοστά SL - 194g έναντι 139 χιλιοστών - 159g ). Και στις δύο δειγματοληψίες του 1997 και 1998 οι δίσκοι είχαν διάφορα μεγέθη. Οι ομάδες ηλικίας διαχωρίστηκαν με τη μέθοδο των γραφικών Petersen ( Bagenal & Tesch , 1978 ). Πολλοί από τους δίσκους στο Uxi Bay έφθασαν σε κατάσταση ετοιμότητας αναπαραγωγής από την αρχή της ανόδου της στάθμης του νερού. Ωστόσο κανένας δίσκος από τους 45 που παρατηρήθηκαν σε galhadas από το lago Urini δύο μήνες νωρίτερα (Οκτώβριος 1997) δεν είχε προχωρήσει σε κατάσταση αναπαραγωγής. Στην κορύφωση της περιόδου των πλημμυρών (τον Απρίλιο του 1999) μόνο ένας από τους 28 δίσκους που ελέγχθηκαν από τα πλημμυρισμένα δάση ήταν σε κατάσταση ετοιμότητας αναπαραγωγής ενώ όλοι οι άλλοι ήταν ανώριμα ή σε κατάσταση ανάπαυσης. Παρατηρήσεις της αναπαραγωγής των δίσκων. Όλα τα κλαδιά που αφαιρέθηκαν από τα galhadas κατά την καταμέτρηση των δίσκων ελέγχθηκαν για αυγά δίσκων αλλά δεν βρέθηκε κανένα. Ένα ζευγάρι των μεγάλων δίσκων με έντονα χρώματα παρατηρήθηκε να γεννάει αυγά σε βυθισμένη ρίζα σε μια θαμνώδεις ακτή του Uxi Bay. Ήταν σε 0,3 - 0,5 μ. βάθος και περίπου 10 μέτρα από την όχθη της λίμνης. Στις τρεις διαδοχικές νύχτες το Δεκέμβριο του 1997 το ζευγάρι ήταν ορατό με το φως του φακού στην ίδια θέση. Κατά τη διάρκεια της ημέρας δεν ήταν ορατό. Μετά από τρεις νύχτες εξαφανίστηκαν, αφήνοντας κάποια γονιμοποιημένα αυγά (διάμετρος 1,3-1,5 mm) στην επιφάνεια της ρίζας. Σε αυτή την περίπτωση προφανώς εγκαταλείφθηκαν. Εντατικές προσπάθειες για τον εντοπισμό άλλων δίσκων να αναπαράγονται ήταν ανεπιτυχείς. Χρώμα δίσκων. Ψηφιακές φωτογραφίες με δίσκους που εξετάστηκαν στο Uxi Bay δεν αποκάλυψαν προφανείς διαφορές στον χρωματισμό ανάμεσα σε αρσενικούς και θηλυκούς δίσκους. Οι μικροί δίσκοι έχουν μια λαδί απόχρωση με σχετικά μικρές περιοχές χρώματος που περιορίζονται στο εδρικό πτερύγιο και στο "κούτελο". Δίσκοι πάνω από 90 mm SL παρουσιάζουν κυματιστές μπλε - πράσινες και κόκκινες γραμμές ή κηλίδες που συνήθως καλύπτουν το μεγαλύτερο μέρος της επιφάνειας του σώματος των ψαριών. Στο εμπόριο ενυδρείο αυθαίρετα ονομάζονται royal όταν οι κυματιστές μπλε - πράσινες και κόκκινες γραμμές εκτείνονται χωρίς διακοπή σε όλο το σώμα του δίσκου. Αυτό τους δίνει μια εντυπωσιακή εμφάνιση και πολύ υψηλότερη εμπορική αξία (Degen , 1995, Crampton , 1999a). 33 από τους 258 (12,8 %) από τα δίσκους που βρέθηκαν σε galhada στο Uxi Bay το 1998 και 13 από τους 24 (54,2 %) των δίσκων που βρέθηκαν σε θαμνώδεις ακτές(σε Uxi Bay και lago Urini) παρουσίαζαν "royal" χρωματισμό . Βρέθηκαν και αρσενικοί και θηλυκοί δίσκοι με χρωματισμό royal με αναλογία 18 αρσενικά με 7 θηλυκά. Μεταξύ των υπόλοιπων δίσκων (που δεν είχαν royal χρωματισμό) εκείνοι που είχαν έρθει σε αναπαραγωγική ετοιμότητα είχαν πιο έντονα χρώματα. Χωρίς κάποια εξαίρεση, ανεξάρτητα από το μέγεθος τους όλοι οι δίσκοι που είχαν παράσιτα (Β. cichlae) είχαν ένα μονότονο χρώμα - παρουσιάζοντας μόνο μικρές υποτονικές περιοχές μπλε - πράσινου ή κόκκινου χρώματος. Οι κάθετες μπάρες στα πλευρά που υποδηλώνουν στρες ή εμφανίζονται στο χαμηλότερα στην ιεραρχία ψάρια υπήρχαν μόνιμα σε αυτούς τους δίσκους. Παρόμοιοι χρωματισμοί παρατηρήθηκαν σε τέσσερις δίσκους που είχαν προσβληθεί με το εντερικό νηματώδες Capillaria.  Παράδειγμα ενός "royal" δίσκου Συζήτηση Διατροφικές προσαρμογές στον υδροβιότοπο του Αμαζονίου. Οι δίσκοι όπως είδαμε προηγουμένως βρέθηκαν να τρέφονται κατά κύριο λόγο με ένα μείγμα αλγών (periphyton), οργανική ύλη και πράσινη φυτική ύλη. Αυτές οι τρεις κατηγορίες συνενώνονται σε μία ενιαία κατηγορία λόγω των πρακτικών δυσκολιών στο διαχωρισμό τους. Η πράσινη φυτική ύλη σε μορφή θρυμματισμένων φύλλων πιθανότατα δε ξεπερνάει το 15 % του συνολικού όγκου (κατ' εκτίμηση). Το υπόλοιπο υλικό ήταν δύσκολο να διαχωριστεί μεταξύ του μερικώς χωνεμένου periphyton και της οργανικής ύλης - η οποία περιλαμβάνει κυρίως φυτικά υπολείμματα που αποσυντίθενται. Το Periphyton περιλαμβάνει πολλών ειδών νηματοειδών αλγών που καλύπτουν τα κλαδιά και τα φύλλα του πλημμυρισμένου δάσους. Είναι μια σημαντική πηγή ενέργειας για τα ψάρια που ζουν σε πλημμυρισμένες όχθες δασών (Araujo - Lima et al , 1986, Forsberg et al 1993). Η οργανική ύλη που παγιδεύεται από το periphyton και συσσωρεύεται στις βυθισμένες επιφάνειες είναι από μόνη της επίσης μια σημαντική πηγή διατροφής για πολλά είδη ψαριών του Αμαζονίου και η κυριότερη πηγή ενέργειας συνολικά για όλα τα είδη των ψαριών (Araujo - Lima et al 1986). Σε περίοδο ξηρασίας υπάρχει πολύ λιγότερο διαθέσιμο βυθισμένο υπόστρωμα για την ανάπτυξη periphyton. Επίσης τα νερά της λίμνης έχουν την τάση να γίνονται θολά από τη δράση των κυμάτων στις όχθες με χώμα με συνέπεια να μειώνεται το φως που βοηθάει την ανάπτυξη των αλγών. Αυτοί οι παράγοντες μπορούν να εξηγήσουν γιατί ένα μεγάλο ποσοστό των δίσκων είχαν μόνο μερικώς γεμάτων στομάχια σε περίοδο ξηρασίας και γιατί τα ασπόνδυλα περιελάμβαναν μεγαλύτερο μέρος της διατροφής τους κατά την περίοδο αυτή. Κατά τη διάρκεια της περιόδου πλημμυρών οι δίσκοι συνήθως βρίσκονται σε καλά φωτισμένα πλημμυρισμένα δάση όπου το periphyton μεγαλώνει πιο πολύ. Σε αυτή την εποχή του έτους τα ασπόνδυλα αποτελούν ένα εκπληκτικά μικρό ογκομετρικά συστατικό της διατροφής τους αν και αποτελούν μια πολύ καλή πηγή ζωτικής πρωτεΐνης για αυτούς. Απαιτείται περαιτέρω μελέτη για τη διερεύνηση της θρεπτικής αξίας των αλγών και των φυτικών και ζωικών υλών στη διατροφή των δίσκων. Το πεπτικό κανάλι του δίσκου αποτελείται από ένα όχι τόσο καλά σχηματισμένο στομάχι και μεγάλο έντερο περίπου 300 mm και πλάτους 3 mm (σε ένα δείγμα μεγέθους 180 mm SL) . Αυτή η εντερική μορφολογία είναι η τυπική στις κιχλίδες κυρίως σε χορτοφάγους , παμφάγους και οργανισμούς που τρέφονται με θρυμματισμένη ύλη. Σαρκοφάγες κιχλίδες όπως οι Cichla και Crenicichla παρουσιάζουν μικρότερα πεπτικά κανάλια με καλά αναπτυγμένα στομάχια (Zihler 1982). Ο Bleher (2006 σελ. . 510-595) αναφέρει λεπτομερείς σημειώσεις με παρατηρήσεις σχετικά με τη διατροφή των δίσκων μετά από πολυετείς παρατηρήσεις. Ανέλυσε το περιεχόμενο των στομάχων πάνω από 8.500 δίσκων και έκανε άμεσες παρατηρήσεις της διατροφής τους στην φύση. Αν και οι περισσότερες από τις διαπιστώσεις του αναφέρονται ποιοτικά ο Bleher παρουσιάζει ορισμένα ποσοτικά στοιχεία για την διατροφή των δίσκων (μολονότι ο αριθμός των δειγμάτων δεν είναι αναφέρεται σελ. 593). Κατά τη διάρκεια των πλημμυρών αναφέρει ότι το μέσο περιεχόμενο του στομάχου έχει: 12 % άλγη και μικροαλγών, 44 % φυτική ύλη (άνθη, καρποί, σπόροι, φύλλα) , 6 % θρυμματισμένη ύλη , 16 % υδρόβια ασπόνδυλα και το 22% χερσαία και δενδρόβια αρθρόποδα. Ενώ κατά τη διάρκεια της ξηρασίας αναφέρει: 25 % άλγη και μικροαλγών, 39 % θρυμματισμένη ύλη, 9 % φυτική ύλη, 22 % υδρόβια ασπόνδυλα και 5 % επίγεια και δενδρόβια αρθρόποδα. Τα δεδομένα συνολικά για όλα τα είδη δίσκων δείχνουν μεγαλύτερο ποσοστό αλγών, φυτικής ύλης και θρυμματισμένης ύλης τόσο κατά τη διάρκεια της ξηρασίας όσο και των πλημμυρών. Τα στοιχεία που παρουσιάζονται εδώ για τους Symphysodon haraldi από την περιοχή Amana δείχνουν μια γενική εικόνα των χαμηλής διατροφικής ποικιλίας και ένα πολύ μεγαλύτερο ποσοστό του periphyton/θρυμματισμένης ύλης από αυτό που αναφέρει ο Bleher. Αυτές οι διαφορές θα μπορούσαν να εξηγηθούν λόγω του μικρού αριθμού των δειγμάτων που αναφέρονται εδώ. Ίσως απαιτούνται πολλά περισσότερα δείγματα για να προσδιοριστεί το πραγματικό εύρος της διατροφής των δίσκων. Δεν αποκλείεται οι διαφορές αυτές να αντανακλούν την φυσική διακύμανση στη διατροφή μεταξύ των διαφορετικών ειδών των δίσκων. Μη τροφικές προσαρμογές στον υδροβιότοπο του Αμαζονίου. Η υπερβολική πλευρική συμπίεση και το ύψους του σώματος του δίσκου είναι κοινά σε μικρότερο βαθμό στα mesonauta και Pterophyllum. Αυτή η μορφολογία είναι προφανώς κατάλληλη για την είσοδο στα στενά διάκενα βυθισμένων κλαδιών. Οι σκούρες κάθετες ρίγες και το επαναλαμβανόμενο πράσινο-καφέ χρώμα των δίσκων παρέχουν προφανώς καμουφλάζ. Οι ραβδώσεις εμφανίζονται και εξαφανίζονται ταχέως ανάλογα με βούληση του ψαριού. Τα πλημμυρισμένα δάση συνήθως έχουν χαμηλό επίπεδο οξυγόνου στο νερό - ακόμα φτάνει σε ανοξικές συνθήκες (σχεδόν καθόλου οξυγόνο στο νερό) καθώς η αποσύνθεση των φύλλων και των οργανικών υπολειμμάτων αφαιρούν το διαθέσιμο οξυγόνο στο νερό (Crampton, 1998). Όπως και άλλες κιχλίδες Αμαζονίου οι δίσκοι παρουσιάζουν μια σειρά από βιοχημικές προσαρμογές που επιτρέπουν την αποτελεσματική αναερόβια αναπνοή κατά τη διάρκεια των περιόδων της υποξίας (έλλειψη οξυγόνου). Τεκμηριώνεται στην μελέτη: Chippari-Gomes et al (2005). Ομαδική συμπεριφορά των δίσκων. Οι δίσκοι σχηματίζουν μεγάλα κοπάδια «αποικίες» στα galhadas τα οποία σε περίοδο ξηρασίας φαίνεται να τα χρησιμοποιούν ως καταφύγια . Οι μικρές αποστάσεις διασποράς που αποδεικνύεται από τα δεδομένα της μελέτης σε συνδυασμό με τον σχετικά μικρό αριθμό των δίσκων που πιάστηκαν σε καινούργια «τεχνητά» galhadas υποδεικνύουν ότι οι αποικίες των δίσκων είναι σχετικά στατικά κατά τη διάρκεια της ξηρασίας. Οι δίσκοι δεν ήταν ομοιόμορφα κατανεμημένοι μεταξύ των galhadas στις περιοχές που μελετήθηκαν. Πολλά φαινομενικά κατάλληλα για αποίκηση galhadas (που βρίσκονταν σε προστατευόμενες τοποθεσίες με 2-4 m βάθος) δεν είχαν καθόλου δίσκους ή είχαν μόνο λίγους ενώ σε άλλα παρόμοια galhadas βρέθηκαν εκατοντάδες δίσκοι. Αυτό δείχνει ότι οι δίσκοι συγκροτούν ενεργά κοινωνικές ομάδες και δεν "συμπληρώνουν" το διαθέσιμο χώρο. Κατά τη διάρκεια της περιόδου των πλημμυρών οι αποικίες των galhadas στο Lago Amana διαλύονται. Οι δίσκοι διασκορπίζονται σε σε γειτονικά πλημμυρισμένα δάση όπου παρατηρήθηκαν μόνοι ή σε μικρές ομάδες (μικρότερες η ίσες των 6 ψαριών). Μελετήθηκε μια αρκετά μεγάλη αποικία στον Uxi Bay και διαπιστώθηκε η επιστροφή ενός ψαριού στην ίδια περιοχή μετά από 11 μήνες πράγμα που δείχνει ότι μερικοί δίσκοι μπορούν να επιστρέφουν συστηματικά στα ίδια καταφύγια κατά τη περίοδο της ξηρασίας (Crampton 1999b), επίσης αναφέρεται η επιστροφή δίσκων σε ίδια galhadas μεταξύ των διαδοχικών περιόδων ξηρασίας σε ένα whitewater ποταμό. Κοπάδιασμα και θήρευση . Ο σχηματισμός μεγάλων κοπαδιών παρατηρείται σε τέτοιο εκτεταμένο βαθμό μόνο στους δίσκους ανάμεσα στις νεο-τροπικές κιχλίδες. Οι περισσότερες κιχλίδες της Νότιας Αμερικής ζουν μόνες ή σε μικρές ομάδες αλλά και σε ζεύγη κατά την αναπαραγωγή και την φροντίδα των αυγών και των γόνων τους (Breder & Rosen 1966, Lowe - McConnell 1969, Goodwin et al 1999). Κοπάδια με περισσότερα από μερικές δεκάδες ψάρια είναι σπάνια σε άλλες κιχλίδες Νότιας Αμερικής (H. Bleher, Η. López - Fernandez, R. Lowe - McConnell). Στην περιοχή Tefé το μόνο είδος κιχλίδας εκτός των δίσκων που έχουν παρατηρηθεί να κοπαδιάζουν (με ενήλικα ψάρια άνω των έξι ατόμων) είναι τα Pterophyllum scalare (Lichtenstein). Αυτό το είδος σχηματίζει ομάδες το πολύ μέχρι 30 ψάρια κατά τη διάρκεια της ξηρασίας(Crampton , 1999c). Κάποιες άλλες κιχλίδες σχηματίζουν ομάδες που δεν υπερβαίνει τα έξι ψάρια όπως οι : C. monoculus, Geophagus proximus, Heros efasciatus, mesonauta mirificus, Uaru amphiacanthoides και Satanoperca jurupari. Θηρευτικά ψάρια μπορούν να φθάσουν σε θεαματικούς αριθμούς κατά τη διάρκεια της ξηρασίας (Goulding 1980) συνεπώς μεγάλα συνεκτικά κοπάδια δίσκων μπορούν να μετριάσουν τη θήρευση. Ο Pitcher (1986) και ο Magurran (1985) παρατήρησαν μηχανισμούς με τους οποίους τα κοπάδια των ψαριών μπορούν να προστατεύουν από θηρευτές. Ο Queiroz & Magurran (2005) πρότεινε ότι το καπάδιασμα του Pygocentrus nattereri ίσως να έχει εξελιχθεί σε απάντηση της θήρευσης. Οι Magurran & Queiroz (2003) σημείωσαν ότι τα κοπάδια του είδους αυτού περιείχαν μεγάλα, ώριμα ψάρια καταλαμβάνοντας κεντρικές θέσεις. Οι Gomez-Laplaza (2002) πρότειναν ότι οι θηρευτές παίζουν σημαντικό ρόλο στο σχηματισμό μικρών κοπαδιών των P. scalare και απέδειξαν ότι η θέση στην ιεραρχία μέσα στο κοπάδι συνδέονταν με τη συμπεριφορά του εκάστοτε θηρευτή. Έχει παρατηρηθεί ότι ψάρια που θα μπορούσαν να τραφούν με δίσκους με βάρος μεγαλύτερο από 200 gr απουσίαζαν από τα galhadas με ομάδες 25 ή περισσοτέρων δίσκων. Ο Bleher (2006 σελ. 503) αναφέρει ότι μερικοί δίσκοι με έντονα χρώματα κολυμπούν έξω από τα galhadas με σκοπό να προστατεύσουν το υπόλοιπο κοπάδι από τα αρπακτικά ψάρια ενεργώντας ως αντιπερισπασμό. Δεν είναι ακόμη σαφές ποια είδη ψαριών είναι σημαντικοί θηρευτές των δίσκων. Τα πιράνχας (Serrasalmus spp, Ν. nattereri) είναι αδηφάγα ιχθυοφάγα αλλά επειδή συνήθως δεν καταπίνουν τη λεία τους η ανάλυση του περιεχόμενου του στομάχου τους μπορεί να αποκρύβει σημαντικά το βαθμό στον οποίο τρώνε δίσκους. Επίσης είναι άγνωστο αν οι δίσκοι βρίσκονται στο διαιτολόγιο (και σε ποιον βαθμό) των caiman, των δελφινιών του ποταμού, των βίδρων, των φιδιών και των πουλιών. Κοπάδιασμα και παράσιτα . Αν και το κοπάδιασμα των δίσκων είναι προσωρινό έχει μερικά μειονεκτήματα. Κυρίως βοηθάει την αύξηση της μετάδοσης νόσων και παρασίτων (Trivers 1985). Ένα μεγάλο ποσοστό (14 %) των δίσκων από τον Uxi Bay είχαν προσβληθεί με το ισόποδο Β. cichlae. Αυτό το παράσιτο όχι μόνο να επιβραδύνει την ανάπτυξη των δίσκων αλλά επίσης περιορίζει την ανάπτυξη ζωηρών χρωμάτων. Το εν λόγω παράσιτο (Braga cichlae) καταβροχθίζει τα βράγχια του ψαριού προκαλώντας μειωμένη πρόσληψη οξυγόνου και πιθανώς εμποδίζοντας έτσι τη φυσιολογική ανάπτυξη. Οι δίσκοι φαίνεται να προσβάλλονται πολύ πιο συχνά από το Β. cichlae από άλλες κιχλίδες που ζουν στον ίδιο βιότοπο παρά το γεγονός ότι δυνητικά προσβάλει οποιοδήποτε ψάρι (Thatcher 1991). Αυτό συμβαίνει διότι οι δίσκοι έχουν πιο έντονο χρώμα από άλλα ψάρια στο περιβάλλον τους και αυτό προσελκύει τα παράσιτα. Υπάρχει ισχυρή θεωρητική βάση που υποστηρίζει ότι το χρώμα χρησιμεύει ως δείκτης για την ποιότητα συντρόφου συμπεριλαμβανομένων της γενικής υγείας και του παρασιτικού φορτίου (Maynard - Smith & Harper 2003). Σύσταση του πληθυσμού . Αν και οι αρσενικοί δίσκοι γίνονται μεγαλύτεροι από τους θηλυκούς και τα δύο φύλα παρουσιάζουν την ίδια αναλογία μήκους και βάρους ενώ η αναλογία αρσενικών/θηλυκών είναι κοντά στο 1:1. Η μελέτη έδειξε ότι το κοπάδι αποτελείται από δύο κύριες ηλικιακές ομάδες μια ομάδα που περιλαμβάνει νέα ψάρια 12 (+ - 6) μηνών και άλλη μία ομάδα με ηλικία 24 (+- 6) μηνών . Επιζώντες μιας 3ης ομάδας η οποία θα περιλαμβάνει τα ψάρια 36 (+- 6) μηνών δεν αποτελούσε εύκολα διακριτή κατηγορία (ήταν ελάχιστα). Οι δίσκοι εκτροφής στα ενυδρεία ζουν συνήθως για 3-5 χρόνια ή πλέον ακόμα περισσότερο (Τ. Silva) αλλά οι άγριοι δίσκοι δείχνουν σύμφωνα με τα στοιχεία να έχουν μικρότερη διάρκεια ζωής. Η ύπαρξη δύο διακριτών ηλικιακών ομάδων στο κοπάδι υποδηλώνει είτε ότι οι δίσκοι δεν παράγουν απογόνους συνεχώς (δεν έχουν πολλαπλές μέσα στη περίοδο των πλημμυρών) είτε ότι η επιβίωση των απογόνων τους δεν είναι σταθερή κατά τη περιόδου των πλημμυρών. Υπάρχουν τρεις υποθέσεις που θα μπορούσαν να το εξηγήσουν. Η πρώτη είναι ότι οι δύο ομάδες προέρχονται από μία μόνο ομάδα των θηλυκών που αναπαράγονται σε δύο διαφορετικές περιπτώσεις. Οι θηλυκοί δίσκοι στα ενυδρεία είναι γνωστό ότι αναπαράγονται τουλάχιστον δύο φορές κάθε χρόνο (Silva & Kotlar 1980). Μετά την ωοτοκία οι δίσκοι ασχολούνται με τη φροντίδα και την υπεράσπιση των γόνων τους για περίπου τρεις έως τέσσερις εβδομάδες (Matthaeus 1992). Μετά το μεγάλωμα της πρώτης γενιάς μπορεί να υπάρχει μια περίοδος κατά την οποία οι γονείς ξεκουράζονται. Τα δεδομένα δείχνουν ότι η αναπαραγωγική ωριμότητα στα galhadas επιτυγχάνεται από τα ψάρια και από τις δύο ηλικιακές ομάδες αλλά αναλογικά πολύ περισσότερο στα ψάρια 2 (+) χρονών. Αυτό το σενάριο όμως είναι απίθανο επειδή η παρατήρηση των πολλαπλών καταστάσεων της αναπαραγωγικής ετοιμότητας των μεγαλύτερων ηλικιακά ψαριών (ομάδα 2 + ετών) κατά την έναρξη των πλημμυρών δεν "δείχνουν" μια συγχρονισμένη ωοτοκία. Μια δεύτερη υπόθεση είναι ότι προέρχονται από διαφορετικές γέννες από τις διάφορες κατηγορίες ηλικίας των θηλυκών. Για παράδειγμα τα 2 + ετών θηλυκά μπορεί να αναπαράγονται νωρίτερα στη σεζόν ενώ τα 1 + ετών θηλυκά μπορεί να αναπαράγονται αργότερα αφού φθάσουν σε πλήρη αναπαραγωγική ετοιμότητα. Ωστόσο η παρατήρηση πολλαπλών καταστάσεων της αναπαραγωγικής ετοιμότητας τόσο στην ομάδα 1 + όσο και στην ομάδα 2 + δείχνει ότι δεν θα μπορούσαν να ξεκινήσουν όλοι αναπαραγωγική δράση την ίδια χρονική στιγμή. Κανονικά αναμένεται τα 2 + ετών θηλυκά να αναπαράγονται κατ' επανάληψη μέσα στη σεζόν και τα 1 + ετών θηλυκά κυρίως προς το τέλος της. Μια τρίτη υπόθεση είναι ότι τα θηλυκά έρχονται σε κατάσταση αναπαραγωγής σύμφωνα με την προηγούμενη περίοδο πλημμυρών (με τα 2 + ετών θηλυκά να κυριαρχούν). Εξωτερικά γεγονότα στον κύκλο των πλημμυρών μπορεί να προκαλούν μια συγχρονισμένη αναπαραγωγή των θηλυκών ή να προκαλέσουν περιόδους αποτυχίας των γενών ή και θάνατο των γόνων τους. * Αυτή η τρίτη υπόθεση αυτή αναλύεται πιο κάτω (βλ. Πότε αναπαράγονται οι δίσκοι). Τοποθεσίες που επιλέγουν οι δίσκοι να γεννήσουν. Τα ζευγάρια δίσκων στα ενυδρεία αποφύγουν τα άλλα ψάρια στο ενυδρείο και αναζητούν προστατευόμενα σημεία στο ενυδρείο για να αναπαραχθούν με σκοπό να μειώσουν τον κανιβαλισμό του γόνου τους (Matthaeus, 1992). Δύο παρατηρήσεις δείχνουν ότι οι δισκοβολία δεν αναπαράγονται στα galhadas. Πρώτον το χαμηλό ποσοστό των έτοιμων δίσκων για αναπαραγωγή στα galhadas σε σχέση με εκείνων στις θαμνώδες ακτές και δεύτερον η πλήρης απουσία των αυγών στα βυθισμένα κλαδιά των galhadas. Στην περιοχή Tefé, πολλές κιχλίδες χρησιμοποιούν φυτά επιφανείας για την προστασία των γόνων τους (π.χ. Α. Nassa, Μ. mirificus και P. scalare) αλλά ενήλικος ή και ανήλικος δίσκος ποτέ δεν βρέθηκε σε τέτοιο το βιότοπο. Η παρατήρηση ενός ζεύγους που φυλούσε τα αυγά του σε μια ρηχή με θάμνους ακτή καθώς και η σύλληψη πολλών δίσκων να είναι σε ετοιμότητα για αναπαραγωγή σε παρόμοιο περιβάλλον υποδηλώνει ότι αυτό το ενδιαίτημα επιλέγουν οι δίσκοι να γεννήσουν. Πότε αναπαράγονται οι δίσκοι. Το μεγαλύτερο μέρος των αναπαραγωγών συνέβησαν στην αρχή της περιόδου των πλημμυρών και όχι νωρίτερα ούτε αργότερα. Αυτό το φαινόμενο είναι κοινό στα ψάρια του Αμαζονίου (Lowe - McConnell 1979, Crampton 1999b) καθώς αυτή την περίοδο διασφαλίζεται ότι οι απόγονοι τους είναι σε θέση να επωφεληθούν από τα άφθονα αποθέματα τροφής και τις χαμηλές πυκνότητες των θηρευτών επειδή διασκορπίζονται στα πλημμυρισμένα δάση αλλά και επίσης επειδή οι γόνοι θα έχουν μεγαλώσει αρκετά κατά την έναρξη της επόμενης σεζόν ξηρασίας όταν οι πυκνότητες των θηρευτών είναι στο υψηλότερο σημείο τους. Αυτό που προκαλεί την έναρξη της αναπαραγωγικής περιόδου στα ψάρια του Αμαζονίου έχουν ήδη μελετηθεί (Kirschbaum 1975, Schwassmann 1978, Lowe - McConnell 1979). Πειράματα που έκανε ο Kirschbaum (1992) απέδειξαν ένα συνδυασμό της μείωσης της αγωγιμότητας του νερού και της φυσικής δράσης της βροχής ως βασικά γεγονότα που προκαλούν την έναρξη της αναπαραγωγής. Οι Kirschbaum & Schugardt (2002) αργότερα απέδειξαν ότι η μείωση της αγωγιμότητας από μόνη της μπορεί να προκαλέσει έναρξη αναπαραγωγής σε ψάρια που ανήκουν σε είδη gymnotiform και mormyriform. Στην περιοχή Tefé η κορύφωση της αναπαραγωγής των δίσκων (Νοέμβριος -Ιανουάριος) αντιστοιχεί στην περίοδο της ανόδου των υδάτων όταν η αγωγιμότητα μειώνεται απότομα. Η περίοδος αυτή δεν συμπίπτει με την περίοδο της μέγιστης βροχόπτωσης στην περιοχή Tefé (Φεβρουάριος-Απρίλιος). Η αρχική άνοδος της στάθμης του νερού του Αμαζονίου και των ποταμών του Japura στην περιοχή Tefé προκαλείται σε μεγάλο βαθμό από τις βροχοπτώσεις στον ανώτερο Αμαζόνιο και όχι από τις τοπικές βροχοπτώσεις (Ayres 1993, Goulding et al 2003). Η ύπαρξη δύο διαφορετικών μεγεθών στη ομάδα 1+ ετών μπορεί να συνδέεται με τα γεγονότα στον κύκλο των πλημμυρών (βλ. τρίτη περίπτωση στη σύσταση του πληθυσμού). Η περιοχή Tefé συνήθως αντιμετωπίζει μία μεγάλη repiquete (παύση της αύξησης της στάθμης του νερού) κατά τη διάρκεια κάθε περιόδου πλημμύρας λόγω του μη συγχρονισμού της ανόδου νερού μεταξύ των ποταμών του Αμαζονίου και του Japura (Ayres 1993). Αυτό το φαινόμενο επηρεάσει την αναπαραγωγή των δίσκων επειδή όπως πολλές κιχλίδες γεννούν σε επιφάνειες κοντά στην επιφάνεια του νερού (Crampton 1999b). Τα αυγά τοποθετούνται κοντά στην επιφάνεια του νερού που είναι καλύτερα οξυγονωμένο και επίσης οι γονείς έχουν να φυλάξουν μικρότερου όγκο νερού από το οποίο θα μπορούσαν να επιτεθούν θηρευτές (Lowe McConnell 1969). Οι αναφορές τοπικών ψαράδων καθώς και προσωπικές παρατηρήσεις δείχνουν ότι πολλές κιχλίδες όπως και οι δίσκοι οι Astronotus ocellatus ( Agassiz ), C. monoculus και Η. efasciatus αποφεύγουν να γεννούν κατά τη διάρκεια που η στάθμη του νερού είναι στάσιμη ή να μειώνεται. Αντίθετα συχνά αναπαράγονται μετά από μερικές ημέρες από τη συνεχή άνοδο της στάθμης του νερού μετά το τέλος του repiquete. Θα εγκαταλείψουν τα αυγά τους σε περίπτωση που υπάρχει ταχεία πτώση στάθμης των υδάτων ή μπορεί να χαθεί όλη η γέννα καθώς θα είναι εκτεθειμένη (Lowe - McConnell 1969). Όλα τα παραπάνω δείχνουν ότι τα μεγάλα σε διάρκεια repiquetes στα τέλη του 1996 και του 1997 θα μπορούσαν να προκαλέσουν μια περίοδο μειωμένων γενών ή και ακόμα περισσότερες αποτυχίες αναπαραγωγής ή και θάνατο των γόνων. Τα repiquetes συνδέονταν με τις διακυμάνσεις στην ηλεκτρική αγωγιμότητα του νερού κατά τη διάρκεια αυτής της μελέτης και οι αλλαγές στην αγωγιμότητα είναι γνωστό ότι είναι το σύνθημα έναρξη της αναπαραγωγικής δραστηριότητας(Kirschbaum & Schugardt 2002). Έτσι οι μεταβολές της αγωγιμότητας μετά από τα repiquetes θα μπορούσαν να προκαλέσουν συγχρονισμένη αναπαραγωγή στους πληθυσμούς των δίσκων. Acknowledgements J. Albert, H. Bleher, T. Hrbek, E. Huusela, H. López Fer- nandez, R. Lowe-McConnell, J. Ready, T. Silva, and S. Willis shared ideas. For field assistance I thank J. Alves de Oliveira and S. Esashika. C. Paxton provided statistical advice. J. Celso-Malta identified B. cichlae. Funding and support were provided by the Mamirauá Institute, Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (grants 380602/96, 2/381597/97-0), and the UK Department for International Development. Funding during the writing stage was provided by National Science Foundation grant DEB- 0614334. Collecting was authorized by IBAMA permits, including 0492/99-12. Animal care followed CNPq guidelines. Literature Cited Araujo-Lima, C. A. R. M., B. Forsberg, R .Victoria & L. Martinelli. 1986. Energy sources for detritivorous fishes in the Amazon. Science, 234: 1256-1258. Araujo-Lima, C. A. R. M. & M. Goulding. 1997. So Fruitful a Fish: Ecology, Conservation and Aquaculture of the Amazon’s Tam- baqui. New York, Columbia University Press. Ayres, J. M. 1993. As Matas de Várzea do Mamirauá - Médio Rio Solimões. Brasília, Brazil, Sociedade Civil Mamirauá/CNPq. Bagenal, T. B. & F. W. Tesch. 1978. Age and growth. Pp. 101-136. In: Bagenal, T (Ed.). Methods for Assessment of Fish Production in Fresh Water. Oxford, Blackwell Scientific Publications. Benedito-Cecilio, E., C. A. R. M. Araujo-Lima, B. R. Forsberg, M. M. Bittencourt & L. A. Martinelli. 2000. Carbon sources of Amazonian fisheries. Fisheries Management and Ecology, 7: 305-314. Bleher, H. 2006. Bleher’s discus. Volume I. Pavia, Italy, Aquapress. Bleher, H., K.N. Stölting, W. Salzburger, & A. Meyer. 2007. Revision of the genus Symphysodon Heckel, 1840 (Teleostei: Perciformes: Cichlidae) based on molecular and morphological characters. Aqua, International Journal of Ichthyology, 12(4): 133-174. Breder, C. M. & D. E. Rosen. 1966. Modes of Reproduction in Fishes. New York, The Natural History Press. Chippari-Gomes, A. R., L. C. Gomes, N. P. Lopes, A. L. Val & V. M. Almeida-Val, 2005. Metabolic adjustments in two Amazonian cichlids exposed to hypoxia and anoxia. Journal of Comparative Physiology, B., 141: 347-355. Correa, S. B., W. G. R. Crampton, L. J. Chapman & J. S. Albert. 2008. Fish assemblages in an upper Amazon floodplain; a com- parison of adjacent habitats during the flood season. Journal of Fish Biology, 72: 629-644. Cox-Fernandes, C. 1997. Lateral migration of fishes in Amazon floodplains. Ecology of Freshwater Fish, 6: 36-44. Crampton, W. G. R. 1998. Effects of anoxia on the distribution, respi- ratory strategies and electric signal diversity of gymnotiform fishes. Journal of Fish Biology, 53 (Supplement A): 307-330. Crampton, W. G. R. 1999a. The impact of the ornamental fish trade on the discus Symphysodon aequifasciatus: a case study from the flood plain forests of Estação Ecológica Mamirauá. Pp. 29-44. In: Padoch, C., J. M. Ayres, M. Pinedo-Vasquez & A. Hender- son (Eds.). Várzea: Diversity, Conservation and Development of Amazonia’s Whitewater Floodplains. New York, New York Botanical Garden Press. Crampton, W. G. R. 1999b. Plano de manejo para o uso sustentável de peixes ornamentais na Reserva Mamirauá. Pp. 159-176. In: Queiroz, H. L., & W. G. R Crampton (Eds.). Estratégias para Manejo de Recursos Pesqueiros em Mamirauá. Brasília, Brazil, Sociedade Civil Mamirauá/CNPq. Crampton, W. G. R. 1999c. Os peixes da Reserva Mamirauá: diver- sidade e história natural na planície alagável da Amazônia. Pp. 10-36. In: Queiroz, H. L. & W. G. R Crampton (Eds.). Estratégias para Manejo de Recursos Pesqueiros em Mamirauá. Brasília, Brazil, Sociedade Civil Mamirauá/CNPq. Degen, B. 1995. Wild-Caught Discus. Neptune City, N.J., T.F.H. Publications. Everhart, W. H., A. W. Eipper & W. D. Young. 1975. Principles of Fishery Science. Ithaca, N.Y., Cornell University Press. Farias, I. P., G. Ortí & A. Meyer. 2000. Total evidence: molecules, morphology, and the phylogenetics of cichlid fishes. Journal of Experimental Zoology, 288:76-92. Farias, I. P, G. Ortí, I. Sampaio, H. Schneider & A. Meyer. 2001. The cytochrome b gene as a phylogenetic marker: the limits of resolution for analyzing relationships among cichlid fishes. Journal of Molecular Evolution, 53:89-103. Farias, I. P., G. Ortí, I. Sampaio, H. Schneider & A. Meyer. 1999. Mi- tochondrial DNA phylogeny of the family Cichlidae: monophyly and fast molecular evolution of the Neotropical assemblage. Journal of Molecular Evolution, 48:703-711. Farias, I. P., H. Schneider & I. Sampaio. 1998. Molecular phylogeny of Neotropical cichlids: the relationships of cichlasomines and heroines. Pp. 499-508. In: Malabarba, L. R., R. R. Reis, R. P. Vari, Z. M. S. Lucena & C. A. S. Lucena (Eds.). Phylogeny and Clas- sification of Neotropical Fishes. Porto Alegre, Brazil, Edipucrs. Forsberg, B. R., C. A. R. M. Araujo-Lima, L. A. Martinelli, R. L. Victoria & J. A. Bonassi. 1993. Autotrophic carbon sources for fish of the Central Amazon. Ecology, 74: 643-652. Frost, W. E. 1943. The natural history of the minnow Phoxinus phoxinus. Journal of Animal Ecology, 12: 139-162. Gomez-Laplaza, L. M. 2002. Social status and investigatory behaviour in the angelfish (Pterophyllum scalare). Behaviour, 139: 1469-1490. Goodwin, N. B., S. Balshine-Earn & J. Reynolds. 1999. Evolutionary transitions in parental care in cichlid fishes. Proceedings of the Royal Society of London, B., 265: 2265-2272. Goulding, M. 1980. The Fishes and the Forest. Berkeley, University of California Press. Goulding, M., R. Barthem & E. Ferreira. 2003. The Smithsonian Atlas of the Amazon. Washington D.C., The Smithsonian Institution. Goulding, M., M. L. Carvalho & E. G. Ferreira. 1988. Rio Negro, Rich Life in Poor Water. Amazonian Diversity and Foodchain Ecology as seen through Fish Communities. The Hague, Netherlands, SPB. Granado-Lorencio, C., C. A. R. M. Araujo-Lima & J. Lobón-Cerviá. 2005. Abundance - distribution relationships in fish assembly of the Amazonas floodplain lakes. Ecography, 28: 515-520. Henderson, P. A., W. D. Hamilton & W. G. R. Crampton. 1998. Evolution and diversity in Amazonian floodplain communities. Pp. 385-419. In: Newbery, D. M., H. H. T. Prins & N. D. Brown (Eds.). Dynamics of Tropical Communities. Oxford, Blackwell Science. Hildemann, W. H. 1959. A cichlid fish Symphysodon discus with unique nurture habits. American Naturalist, 93: 27-34. Hyslop, E. J. 1980. Stomach contents analysis - a review of methods and their application. Journal of Fish Biology, 17: 411-429. Jearld, J. A. 1983. Age determination. Pp. 301-324. In: Nielsen, L.A., & D. L. Johnson (Eds.). Fisheries Techniques. Bethesda, MD, American Fisheries Society. Jepsen, D. B., K. O. Winemiller & D. C. Taphorn. 1997. Temporal patterns of resource partitioning among Cichla species in a Vene- zuelan blackwater river. Journal of Fish Biology, 51: 1085-1108. Jepsen, D. B., K. O. Winemiller, D. C. Taphorn & D. R. Olarte. 1999. Age structure and growth of peacock cichlids from rivers and reservoirs of Venezuela. Journal of Fish Biology, 55: 433-450. Junk, W. J. 1997. General aspects of floodplain ecology with special reference to Amazonian floodplains. Pp. 3-20. In: Junk, W. J. (Ed.). The Central Amazon floodplain: ecology of a pulsing system. Berlin, Springer-Verlag. Kirschbaum, F. 1975. Environmental factors control the periodical reproduction of tropical electric fish. Experientia, 31: 1159-1160. Kirschbaum, F. 1992. Cyclic reproduction of tropical freshwater fishes: comparative experimental aspects. Pp. 115-123. In: Adamek, Z. & M. Flajshans (Eds.). Fish Reproduction 1992, Conference Proceedings. Vodnany, Czechoslovakia. Kirschbaum, F., & C. Schugardt. 2002. Reproductive strategies and developmental aspects in mormyrid and gymnotiform fishes. Journal of Physiology-Paris, 96:557-566. Kullander, S. O. 1986. The Cichlid Fishes of the Amazon River Drai- nage of Peru. Stockholm, Swedish Museum of Natural History. Kullander, S. O. 1996. Eine weitere Ubersicht der Diskusfische, Gattung Symphysodon Heckel. Pp. 10-16. In: R. Stawikowski (Ed.). DATZ Sonderheft Diskus. Verlag Eugen Ulmer, Stuttgart. Kullander, S.O. 1998. A phylogeny and classification of the South American Cichlidae (Teleostei: Perciformes). Pp. 461-498. In: L. R. Malabarba, R. R. Reis, R. P. Vari, Z. M. S. Lucena and C. A. S. Lucena (Eds.). Phylogeny and Classification of Neotropical Fishes. Porto Alegre, Brazil, Edipucrs. Lowe-McConnell, R. H. 1969. The cichlid fishes of Guyana, South America, with notes on their ecology and breeding behaviour. Zoological Journal of the Linnean Society, 48: 255-302. Lowe-McConnell, R. H. 1979. Ecological aspects of seasonality in fishes of tropical waters. Symposia of the Zoological Society of London, 44: 219-241. Lundberg, J. G., L. G. Marshall, J. Guerrero, B. Horton, M. C. S. L. Malabarba & F. Wesselingh. 1998. The Stage for Neotropical Fish Diversification: A History of Tropical South American Rivers. Pp. 13-48. In: L. R. Malabarba, R. R. Reis, R. P. Vari, Z. M. S. Lucena, and C. A. S. Lucena (Eds.). Phylogeny and Classification of Neotropical Fishes. Edipucrs, Porto Alegre. Lyons, E. 1959. Symphysodon discus tarzoo. New blue discus elec- trify aquarium world. Tropicals Magazine, 4: 6-8,10. Magurran, A. E., W. Oulton & T. J. Pitcher. 1985. Vigilant behaviour and shoal size in minnows. Zeitschrift für Tierpyschologie, 67: 167-178. Magurran, A. E. & H. L. Queiroz. 2003. Partner choice in piranha shoals. Behaviour, 140: 289-299. Matthaeus, W. 1992. Observations on the behaviour of Symphysodon aequifasciatus. Freshwater and Marine Aquarium, 15: 12-16. Mayland, H. J. 1994. Adventures with Discus. Neptune City, N.J., T.F.H. Publications. Maynard Smith, J. & D. Harper. 2003. Animal Signals. Oxford, Oxford University Press. Mesquita, D. R., J. I. R. Porto & E. Feldberg. 2008. Chromosomal varia- bility in the wild ornamental species of Symphysodon (Perciformes: Cichlidae) from Amazon. Neotropical Ichthyology, 6(2): 181-190. Nikolsky, G. V. 1963. The Ecology of Fishes. Academic Press, London. Petry, P., P. B. Bayley & D. F. Markle. 2003. Relationships between fish assemblages, macrophytes and environmental gradients in the Amazon River floodplain. Journal of Fish Biology, 63: 547-579. Pitcher, T. J. 1986. Functions of shoaling behaviour in teleosts. Pp. 294-338. In: Pitcher, T. J. (Ed.). The Behaviour of Teleost Fishes. London, Croom Helm. Queiroz, H. L. & A. Magurran. 2005. Safety in numbers? Shoaling behaviour of the Amazonian red-bellied piranha. Biology Letters, 1: 155-157. Ready, J. S., E. J. G. Ferreira & S. O. Kullander. 2006. Discus fishes: mitochondrial DNA evidence for a phylogeographic barrier in the Amazon genus Symphysodon (Teleostei: Cichlidae). Journal of Fish Biology, 69 (Supplement B): 200-211. Schultz, L. P. 1960. A review of the pompadour or discus fishes, genus Symphysodon of South America. Tropical Fish Hobbyist, 8: 5-17. Schwassmann, H. O. 1978. Times of annual spawning and reproduc- tive strategies in Amazonian fishes. Pp. 187-200. In: Thorpe, J.E. (Ed.). Rhythmic Activity of Fishes. London, Academic Press. Silva, T. & B. Kotlar. 1980. Discus. Neptune City, N.J., T.F.H. Publications. Southwood, T. R. E., & Henderson, P. A. 2000. Ecological Methods. Third Edition. Oxford, Blackwell Science. Swynnerton, G. H. & E. B. Worthington. 1940. Notes on the food of fish in Haweswater (Westmorland). Journal of Animal Ecology, 9: 183-187. Thatcher, V. E. 1991. Amazon fish parasites. Amazoniana, 11: 236-572. Trivers, R. L. 1985. Social Evolution. Menlo Park, CA., Benjamin- Cummings. Val, A. L. & V. M. F. Almeida-Val. 1995. Fishes of the Amazon and their Environment. New York, Springer. Winemiller, K. O., D. C. Taphorn & A. Barbarino-Duque. 1997. Eco- logy of Cichla (Cichlidae) in two blackwater rivers of southern Venezuela. Copeia, 1997: 690-696. Zihler, F. 1982. Gross morphology and configuration of digestive tracts of Cichlidae (Teleostei, Perciformes): phylogenetic and functional significance. Netherland Journal of Zoology, 32: 544-571. Accepted July 2008 Published December 22, 2008
__________________
Gab at Facebook: https://www.facebook.com/GreekAquaristsBoards/ https://www.facebook.com/groups/GreekAquaristsBoards/ Knowledge is having the right answer Intelligence is asking the right question Τελευταία επεξεργασία από το χρήστη Γιώργος Κ : 11-03-14 στις 20:16 
|
|
#2
11-03-14, 00:08
|
||||
|
||||
|
Φοβερη δουλεια.
Τωρα πρεπει να παρουμε και μια βδομαδα αδεια για να τη διαβασουμε προσεκτικα
__________________
O delta66 δεν εχει καμια σχεση με επαγγελματα που αφορουν το ενυδρειο και δεν εξαρταται απο σπονσορες, επαγγελματιες και διαφημισεις.
Γραφει ελευθερα και ανεπηρεαστα την αποψη του και ευχαριστει το GAB που του δινει αυτη τη δυνατοτητα.
|
|
#4
11-03-14, 01:24
|
||||
|
||||
|
Πολύ καλή προσπάθεια Γιώργο... θαυμάζω το κουράγιο σου.
 Χρειάζονται κάποιες διορθώσεις της μετάφρασης... για να γίνει πιο σωστό το κείμενο.-
|
|
#5
11-03-14, 01:53
|
||||
|
||||
|
Παράθεση:
Παράθεση:
Παράθεση:
Παράθεση:
__________________
Gab at Facebook: https://www.facebook.com/GreekAquaristsBoards/ https://www.facebook.com/groups/GreekAquaristsBoards/ Knowledge is having the right answer Intelligence is asking the right question
|
|
#6
11-03-14, 10:11
|
||||
|
||||
|
Σ' ευχαριστούμε ακάματε δισκοπαθή!
|
|
#7
11-03-14, 11:29
|
||||
|
||||
|
Γιωργο αψογος οπως παντα, Μπραβο
|
|
#8
11-03-14, 11:35
|
||||
|
||||
|
Πολύ καλή δουλειά.
Ασχετος με τους δίσκους αλλά κάτι τέτοια άρθρα πιστεύω πως αξίζει το κόπο να τα διαβάσει κάποιος. Ευχαριστούμε Γιώργο (Ακόμα το διαβάζω  ) )
__________________
Δεν απαντώ σε πμ για ενυδρειακά θέματα Φυλλάδιο GAB - Χρυσόψαρα Φυλλάδιο GAB - Μονομάχοι Τα ενυδρεία μου 
|
|
#9
12-03-14, 19:47
|
||||
|
||||
|
μπραβο ρε γιωργο πολυ καλο
__________________
Ένας βλάκας που περπατάει, αξίζει πιο πολύ από δέκα διανοούμενους που κάθονται...
|
|
#15
12-03-14, 22:39
|
||||
|
||||
|
Πολυ καλο,και γω στο διαβασμα ειμαι.(Γιατι να το κρυψωμεν αλλωστε)
Ανέλυσε το περιεχόμενο των στομάχων πάνω από 8.500 δίσκων και έκανε άμεσες παρατηρήσεις της διατροφής τους στην φύση. Αν και οι περισσότερες από τις διαπιστώσεις του αναφέρονται ποιοτικά ο Bleher παρουσιάζει ορισμένα ποσοτικά στοιχεία για την διατροφή των δίσκων (μολονότι ο αριθμός των δειγμάτων δεν είναι αναφέρεται σελ. 593). Κατά τη διάρκεια των πλημμυρών αναφέρει ότι το μέσο περιεχόμενο του στομάχου έχει: 12 % άλγη και μικροαλγών, 44 % φυτική ύλη (άνθη, καρποί, σπόροι, φύλλα) , 6 % θρυμματισμένη ύλη , 16 % υδρόβια ασπόνδυλα και το 22% χερσαία και δενδρόβια αρθρόποδα. Ενώ κατά τη διάρκεια της ξηρασίας αναφέρει: 25 % άλγη και μικροαλγών, 39 % θρυμματισμένη ύλη, 9 % φυτική ύλη, 22 % υδρόβια ασπόνδυλα και 5 % επίγεια και δενδρόβια αρθρόποδα. Τα δεδομένα συνολικά για όλα τα είδη δίσκων δείχνουν μεγαλύτερο ποσοστό αλγών, φυτικής ύλης και θρυμματισμένης ύλης τόσο κατά τη διάρκεια της ξηρασίας όσο και των πλημμυρών. Τα στοιχεία που παρουσιάζονται εδώ για τους Symphysodon haraldi από την περιοχή Amana δείχνουν μια γενική εικόνα των χαμηλής διατροφικής ποικιλίας και ένα πολύ μεγαλύτερο ποσοστό του periphyton/θρυμματισμένης ύλης από αυτό που αναφέρει ο Bleher. Αυτές οι διαφορές θα μπορούσαν να εξηγηθούν λόγω του μικρού αριθμού των δειγμάτων που αναφέρονται εδώ. Ίσως απαιτούνται πολλά περισσότερα δείγματα για να προσδιοριστεί το πραγματικό εύρος της διατροφής των δίσκων. Δεν αποκλείεται οι διαφορές αυτές να αντανακλούν την φυσική διακύμανση στη διατροφή μεταξύ των διαφορετικών ειδών των δίσκων. Αυτο μας λεει πως ειναι κυριος χορτοφαγοι ")
|
|
#16
13-03-14, 00:25
|
||||
|
||||
|
Βασικα τους κατατασσει σαν παμφαγους-χορτοφαγους οργανισμους:
Παράθεση:
Παράθεση:
δηλαδη ενω θα μπορουσαν να φανε πρωτεΐνη σε εποχη που ειναι πιο προσβασιμη ακομα και τοτε επιλεγουν να τρωνε φυτικες τροφες ως επι το πλειστον!! Θεωρω οτι εχουν ιδιο το πεπτικο συστημα και οι δισκοι στα ενυδρεια (ακομα και οι F10000) αρα αναρωτιεμαι και με προβληματιζει αν τους "αναγκαζουμε" να τρωνε υπερβολικη πρωτεινη ενω δε θα επρεπε!
__________________
Gab at Facebook: https://www.facebook.com/GreekAquaristsBoards/ https://www.facebook.com/groups/GreekAquaristsBoards/ Knowledge is having the right answer Intelligence is asking the right question
|
|
#17
13-03-14, 21:00
|
||||
|
||||
|
1. Γιώργο... θέλει και κάποιες διορθώσεις στο συντακτικό. Θα κανονίσουμε... να το δούμε παρέα.
2. Κάποιοι... ανακαλύπτουν τον τροχό; Ο Bleher έχει πει (στα σεμινάρια)... ότι στα στομάχια των δίσκων ΔΕΝ έχει βρει κρέας. Απ' την άλλη ακούγετε και αυτό το (κλασικό πλέον)... οι δίσκοι ΔΕΝ ανοίγουν τις κοιλιές των αγελάδων στον Αμαζόνιο... για να τους φάνε τις καρδιές. Νομίζω ότι ΑΝ ανοίξει αυτή η κουβέντα... θα δημιουργηθεί φαύλος κύκλος. Όποιος θέλει να δίνει μπανάνες στους δίσκους του ας το κάνει... κι' ας ελπίσει να τις τρώνε.-
|
|
#18
13-03-14, 21:41
|
||||
|
||||
|
Τορα, με αυτα που διαβαζουμε, και με αυτα που ειπε στα σεμιναρια ο Bleher δεν μπορουμε να πουμε, οτι ο τροχος ειναι στογγυλος??
|
|
#19
13-03-14, 21:43
|
||||
|
||||
|
Είναι;
|
|
#20
13-03-14, 22:10
|
||||
|
||||
|
Εγω λεω οτι ειναι
|
|
#21
13-03-14, 23:06
|
||||
|
||||
|
Εγώ... ακόμα το ψάχνω!!!!!
|
|
#23
28-10-16, 11:59
|
||||
|
||||
|
Mερικές φωτογραφίες του φυσικού περιβάλλοντος στο οποίο ζούν οι δίσκοι.
__________________

|
|
#24
28-10-16, 13:08
|
||||
|
||||
|
Ενδιαφερουσες οι φωτογραφίες, ωραίος Δημήτρη. Ενδιαφέρον φάνηκε και το άρθρο Γιώργο, θα κάτσω να το διαβάσω ένα απόγευμα.
|
|
#25
28-10-16, 15:54
|
||||
|
||||
|
Ωραίος Δημήτρη!!!!!
Γιατί ΔΕΝ είδα... καρδινάλιους, ροδόστομους, αρλεκίνους κλπ. ;
|
|
#26
30-07-17, 13:23
|
||||
|
||||
|
'Eνα ενδιαφέρον βίντεο λίγο διαφορετικό απο τα υπόλοιπα που κυκλοφορούν σχετικά με το περιβάλλον που ζούν οι άγριοι δίσκοι.
Βλέπουμε οτι στο σημείο αυτό του ποταμού υπάρχει αυξημένη ροή του νερού.
__________________
|
|
| Συνδεδεμένοι χρήστες που διαβάζουν αυτό το θέμα: 1 (0 μέλη και 1 επισκέπτες) | |
|
|
 Παρόμοια Θέματα
Παρόμοια Θέματα
|
||||
| Θέμα | Δημιουργός | Forum | Απαντήσεις | Τελευταίο Μήνυμα |
| Για δίσκους, γυαλες, χομπυ, την φύση και τα ζώα... | drbig | Γλυκό νερό - Γενικά | 165 | 17-09-10 12:37 |
| Συγκάτοικοι για τους δίσκους μου... | Theokyr | Δίσκοι | 19 | 04-06-06 20:05 |
| Βοήθεια χάνω τους δίσκους !!!! | ΚΩΣΤΑΣ ΚΕΦΑΛΑΣ | Δίσκοι | 24 | 09-02-05 10:46 |
| Χορεύοντας με τους Δίσκους | jumpy | Δίσκοι | 5 | 18-01-03 19:01 |
|





















 Αλλαγή σε γραμμικό τρόπο
Αλλαγή σε γραμμικό τρόπο


